WikiDer > Rsa RNK
Rsa RNKlari bor kodlamaydigan RNKlar bakteriyada topilgan Staphylococcus aureus. Umumiy ism ularning kashfiyotidan kelib chiqadi va buni anglatmaydi homologiya. Bioinformatika skanerlash natijasida RsaA-K va RsaOA-OG nomli 16 Rsa RNK oilalari aniqlandi.[1][2] Boshqalar, RsaOH-OX, RNomic yondashuvi tufayli topilgan.[3] RNKlarda turli xil ekspression naqshlari ko'rsatilgan bo'lsa-da, yangi kashf etilgan RNKlarning aksariyati ko'rsatildi Hfq- mustaqil va ko'pi bilan olib boriladigan a C- boy motif (UCCC).[1]
RsaA
MgrA transkripsiya regulyatorining mRNK bilan bog'lanishini tarjima qilishni bostiradi, biofilm shakllanishini kuchaytiradi va bakteriyalarning virusliligini pasaytiradi.[4] Boshqa mRNAlar: shu jumladan peptidoglikan metabolizmasida ishtirok etadigan SsaA o'xshash fermentlar va salgılanan yallig'lanishga qarshi FLIPr oqsillari RsaA ning to'g'ridan-to'g'ri maqsadlari sifatida tasdiqlangan.[5]
RsaE
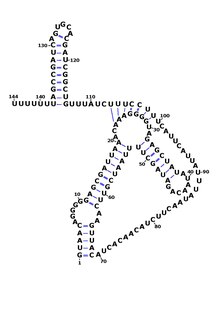
RsaE jinsning boshqa a'zolarida uchraydi Stafilokokk kabi Staphylococcus epidermidis va Staphylococcus saprophyticus va ushbu turdan tashqarida topilgan yagona Rsa RNKidir Makrokokus caseolyticus va Bacillus. Yilda Bacillus subtilis, RsaE ilgari ncr22 sifatida aniqlangan edi.[8][9] RsaE doimiy ravishda quyi oqimda joylashgan PepF qaysi kodlar uchun oligoendopeptidaza F. yordamida RsaE funktsiyasi aniqlandi genlarni nokaut qilish tahlil va genning haddan tashqari ekspressioni - bu tartibga solinishi aniqlandi ifoda ishtirok etgan bir qancha fermentlarning metabolizm orqali antisens ularning majburiyligi mRNA.[1][3]
RsaE azot oksidi (NO) borligi bilan tartibga solinishi ko'rsatilgan. Yilda Bacillus subtilis u oksidlanish stresi va oksidlanish-qaytarilish reaktsiyalari bilan bog'liq funktsiyalar bilan genlarning ekspressionini boshqaradi va u RoxS (oksidlovchi stress bilan bog'liq) deb o'zgartirildi.[10]
RsaF
Yilda S.aureus RsaF turlari RsaE bilan bir xil intergenik mintaqada joylashgan va RsaE ning 3 ′ uchi bilan 20bp ga teng. RsaE dan farqli o'laroq, RsaF va uning yuqori oqimdagi geni faqat aniqlangan S.aureus turlari.[1]
RsaK
RsaK topilgan etakchining ketma-ketligi ning glcA anrni kodlaydigan mRNA ferment glyukozaga xos ishtirok etadi fosfotransferaza tizimi. RsaK tarkibida konservalangan ribonukleik ham mavjud antiterminator GclT oqsili tan olgan tizim.[11]
RsaI
RsaOG[2] shuningdek, RsaI deb o'zgartirildi[1] ichida toksin yoki invaziya mexanizmlarini boshqarishni aniq sozlaydi deb o'ylashadi S. aureus orqali trans-aktyorlik mexanizmlar. Uning ikkilamchi tuzilish o'z ichiga oladi pseudoknot ikkita yuqori konservalangan juft bo'lmagan ketma-ketliklar o'rtasida hosil bo'lgan.[6]
Ifoda naqshlari
RsaD, E H va men yuqori darajada ifodalanganligi aniqlandi S. aureus. Boshqa Rsa RNKlarining ifoda darajasi har xil atrof-muhit sharoitida o'zgarib turardi, masalan RsaC tomonidan induktsiya qilingan sovuq zarba va RsaA javoban induktsiya qilinadi ozmotik stress.[1][2][3]
RsaE va RsaF genlari bir-biriga to'g'ri keladi S.aureus turlar, ammo qarama-qarshi ifoda naqshlariga ega ko'rinadi.[1] Transkripsiya aralashuvi a o'rtasidagi o'zaro bog'liqlik tufayli σA tanib olish motifi va potentsial σB majburiy sayt ikkita transkriptning differentsial ifodasini keltirib chiqaradigan mexanizm sifatida taklif qilingan[1][12]
Shuningdek qarang
Adabiyotlar
- ^ a b v d e f g h Geissmann T, Chevalier C, Cros MJ, Boisset S, Fechter P, Noirot C, Schrenzel J, Francois P, Vandenesch F, Gaspin C, Romby P (Noyabr 2009). "Staphylococcus aureus-da kichik kodlamaydigan RNKlarni qidirishda regulyatsiya uchun saqlanib qolgan ketma-ketlik motifi aniqlandi". Nuklein kislotalarni tadqiq qilish. 37 (21): 7239–7257. doi:10.1093 / nar / gkp668. PMC 2790875. PMID 19786493.
- ^ a b v Marchais A, Naville M, Bohn C, Bouloc P, Gautheret D (iyun 2009). "Filogenetik profillar yordamida bakterial genomdagi barcha kodlash ketma-ketliklarining bir martalik tasnifi". Genom tadqiqotlari. 19 (6): 1084–1092. doi:10.1101 / gr.089714.108. PMC 2694484. PMID 19237465.
- ^ a b v Bohn C, Rigoulay C, Chabelskaya S, Sharma CM, Marchais A, Skorski P, Borezee-Durant E, Barbet R, Jacquet E, Jacq A, Gautheret D, Felden B, Vogel J, Bouloc P (oktyabr 2010). "Staphylococcus aureus'dagi kichik RNKlarning eksperimental kashfiyoti natijasida markaziy metabolizmning riboregulyatori aniqlandi". Nuklein kislotalarni tadqiq qilish. 38 (19): 6620–6636. doi:10.1093 / nar / gkq462. PMC 2965222. PMID 20511587.
- ^ Romilly C, Lays C, Tomasini A, Caldelari I, Benito Y, Hammann P, Geissmann T, Boisset S, Romby P, Vandenesch F (mart 2014). "Kodlamaydigan RNK Staphylococcus aureus'dagi regulyatorni boshqarish orqali bakteriyalarning turg'unligini kuchaytiradi va virulentlikni pasaytiradi". PLoS patogenlari. 10 (3): e1003979. doi:10.1371 / journal.ppat.1003979. PMC 3961350. PMID 24651379.
- ^ Tomasini A, Moreau K, Chicher J, Geissmann T, Vandenesch F, Romby P, Marzi S, Caldelari I (iyun 2017). "Staphylococcus aureus kodlamaydigan RNK RSAA ning RNK-ob'ekti: hujayra yuzasi xususiyatlari va himoya mexanizmlariga ta'siri". Nuklein kislotalarni tadqiq qilish. 45 (11): 6746–6760. doi:10.1093 / nar / gkx219. PMC 5499838. PMID 28379505.
- ^ a b Marchais A, Bohn C, Bouloc P, Gautheret D (mart 2010). "RsaOG, yangi stafilokokklar oilasi, yuqori transkripsiya qilingan kodlanmaydigan RNK". RNK biologiyasi. 7 (2): 116–119. doi:10.4161 / rna.7.2.10925. PMID 20200491.
- ^ Darti K, Denis A, Ponti Y (avgust 2009). "VARNA: interfaol chizish va RNK ikkilamchi tuzilishini tahrirlash". Bioinformatika. 25 (15): 1974–1975. doi:10.1093 / bioinformatika / btp250. PMC 2712331. PMID 19398448.
- ^ Rasmussen S, Nilsen XB, Jarmer H (sentyabr 2009). "Bacillus subtilis genomidagi transkripsiyaviy faol hududlar". Molekulyar mikrobiologiya. 73 (6): 1043–1057. doi:10.1111 / j.1365-2958.2009.06830.x. PMC 2784878. PMID 19682248.
- ^ Irnov I, Sharma CM, Vogel J, Winkler WC (oktyabr 2010). "Bacillus subtilis-da regulyativ RNKlarni aniqlash". Nuklein kislotalarni tadqiq qilish. 38 (19): 6637–6651. doi:10.1093 / nar / gkq454. PMC 2965217. PMID 20525796.
- ^ Durand S, Braun F, Lioliou E, Romilly C, Helfer AC, Kuhn L, Quittot N, Nikolas P, Romby P, Condon C (fevral, 2015). "Azot oksidi bilan boshqariladigan kichik RNK Bacillus subtilisdagi oksidlanish-qaytarilish gomeostaziga aloqador genlarning ekspresiyasini boshqaradi". PLoS Genetika. 11 (2): e1004957. doi:10.1371 / journal.pgen.1004957. PMC 4409812. PMID 25643072.
- ^ Langbein I, Bachem S, Stulke J (1999 yil noyabr). "Bacillus subtilis transkripsiya antiterminatori GlcT ning RNK bilan bog'lanish sohasining uning RNK maqsadi, RAT bilan o'ziga xos o'zaro ta'siri". Molekulyar biologiya jurnali. 293 (4): 795–805. doi:10.1006 / jmbi.1999.3176. PMID 10543968.
- ^ Shearwin KE, Kallen BP, Egan JB (iyun 2005). "Transkripsiya aralashuvi - avariya kursi". Genetika tendentsiyalari. 21 (6): 339–345. doi:10.1016 / j.tig.2005.04.009. PMC 2941638. PMID 15922833.
Qo'shimcha o'qish
- Pichon C, Felden B (oktyabr 2005). "Staphylococcus aureus genomik va patogenlik orollaridan ma'lum bo'lgan kichik RNK genlari patogen shtammlar orasida o'ziga xos ifoda bilan". Amerika Qo'shma Shtatlari Milliy Fanlar Akademiyasi materiallari. 102 (40): 14249–14254. doi:10.1073 / pnas.0503838102. PMC 1242290. PMID 16183745.
- Altuvia S (2007 yil iyun). "Bakterial kichik kodlamaydigan RNKlarni aniqlash: eksperimental yondashuvlar". Mikrobiologiyaning hozirgi fikri. 10 (3): 257–261. doi:10.1016 / j.mib.2007.05.003. PMID 17553733.
- Morfeldt E, Teylor D, fon Gabeyn A, Arvidson S (sentyabr 1995). "Stafilokokk aureusida alfa-toksinli tarjimani transkodlangan antisensli RNK, RNAIII tomonidan faollashtirish". EMBO jurnali. 14 (18): 4569–4577. doi:10.1002 / j.1460-2075.1995.tb00136.x. PMC 394549. PMID 7556100.
- Roberts C, Anderson KL, Murphy E, Projan SJ, Mount V, Hurlburt B, Smeltzer M, Overbeek R, Disz T, Dunman PM (aprel 2006). "Staphylococcus aureus virulence factor regulator, SarA ning log-faza mRNA yarim umrlariga ta'sirini tavsiflovchi". Bakteriologiya jurnali. 188 (7): 2593–2603. doi:10.1128 / JB.188.7.2593-2603.2006. PMC 1428411. PMID 16547047.
- Anderson KL, Roberts C, Disz T, Vonstein V, Xvan K, Overbek R, Olson PD, Projan SJ, Dunman PM (oktyabr 2006). "Staphylococcus aureus issiqlik zarbasi, sovuq zarba, qattiq va SOS reaktsiyalarining xarakteristikasi va ularning log-fazali mRNA aylanishiga ta'siri". Bakteriologiya jurnali. 188 (19): 6739–6756. doi:10.1128 / JB.00609-06. PMC 1595530. PMID 16980476.
- Huntzinger E, Boisset S, Saveanu C, Benito Y, Geissmann T, Namane A, Lina G, Etienne J, Ehresmann B, Ehresmann C, Jakier A, Vandenesch F, Romby P (fevral 2005). "Staphylococcus aureus RNAIII va endoribonuklease III kurort genlarining ekspressionini muvofiqlashtiradi". EMBO jurnali. 24 (4): 824–835. doi:10.1038 / sj.emboj.7600572. PMC 549626. PMID 15678100.
- Novick RP, Ross HF, Projan SJ, Kornblum J, Kreiswirth B, Moghazeh S (oktyabr 1993). "Stafilokokklar virulentligi omillarini sintezi tartibga soluvchi RNK molekulasi tomonidan boshqariladi". EMBO jurnali. 12 (10): 3967–3975. doi:10.1002 / j.1460-2075.1993.tb06074.x. PMC 413679. PMID 7691599.
- Bohn C, Rigoulay C, Bouloc P (2007 yil fevral). "Aureus Staphylococcus da RNK bilan bog'langan Hfq oqsilining aniqlanadigan ta'siri yo'q". BMC mikrobiologiyasi. 7: 10. doi:10.1186/1471-2180-7-10. PMC 1800855. PMID 17291347.
Rsa RNK ikkilamchi tuzilish rasmlari galereyasi | |
|---|---|
|









